

Kartlegging av AH1-mutasjonen i NRF
AH1 er ein recessiv mutasjon som har gitt problem med fruktbarheit og kalvehelse i ayrshire-rasen.
Avlsforskar i Geno
arne.gjuvsland@geno.no
Avlsforskar i Geno
marte.holtsmark@geno.no



AH1-mutasjonen har gitt problem med fruktbarheit og kalvehelse hos Ayrshire blant anna i Finland. I NRF er frekvensen så låg at mutasjonen i dag er eit lite problem.
Foto:iStockphoto
Med innføringa av genomisk seleksjon verda over blir stadig fleire dyr genotypa. Hovudformålet med genotyping av dyr er å rekna genomiske avlsverdiar, men genotypane som blir samla inn kan også brukast til andre formål som å kontrollera slektskap eller kartlegga mutasjonar med stor effekt på viktige eigenskapar. I Buskap nummer 2 i 2018 skreiv me generelt om recessive mutasjonar med negativ effekt på fruktbarheit og her går me nærare inn på ein slik mutasjon kalt AH1. AH1 har gitt problem med fruktbarheit og kalvehelse i ayrshire-rasen i Nord-Amerika og Finland, og me har no gjort ei kartlegging av mutasjonen i NRF.
AH1 i Ayrshire i USA og Canada
Den første rapporten om AH1 var frå ayrshire-populasjonen i USA. Ein artikkel i Journal of Dairy Science i 2014 rapporterte om eit område på 300 000 basepar på kromosom 17 som inneheldt ein recessiv mutasjon med negativ effekt på fruktbarheit og kalla den Ayrshire Haplotype 1 (AH1). Mutasjonen blei spora tilbake til oksen Selwood Betty’s Commander (fødd 1953), den mest brukte oksen i ayrshire-rasen i USA. Etterkommarane hans var også svært populære, og dette førte til at frekvensen av AH1 i USA auka raskt og har vore ganske stabil sidan rundt 1970. Når mutasjonen blei oppdaga i 2014 var bærarfrekvensen i USA 26 prosent. Den første studien fann at oksar som var bærarar av AH1 hadde fleire insemineringar per kalv, men hadde for lite datagrunnlag til å kunne konkludera om effekt på dødfødslar. Ei ny kartlegging i canadisk Ayrshire, der bærarfrekvensen også er over 20 prosent, avdekka ein auke i dødfødslar på to prosentpoeng dersom både far og morfar til kalven var AH1-bærarar. Med dei høge bærarfrekvensane i USA og Canada er AH1 ei viktig årsak til redusert fruktbarheit i ayrshire-rasen, men å utelukka bærarar som avlsoksar vil føra til tapt avlsframgang. Derfor er det sett i gang tiltak for å unngå bærar-bærar inseminasjonar, mens ein over tid fjernar AH1 frå populasjonen. I studien i USA blei det ikkje gjort forsøk på å finna den nøyaktige posisjonen til mutasjonen, men dette blei gjort like etterpå i ein studie i Finsk Ayrshire.
AH1-status far |
AH1-status morfar |
Tal på kalvingar |
Dødfødslar |
|---|---|---|---|
Ikkje-bærar |
Ikkje-bærar |
473 298 |
3,74 prosent |
Ikkje-bærar |
Bærar |
11 518 |
3,57 prosent |
Bærar |
Ikkje-bærar |
35 492 |
3,63 prosent |
Bærar |
Bærar |
1 226 |
5,22 prosent |
AH1 og PIRM i Finsk Ayrshire
I perioden 2011-2014 rapporterte finske mjølkebønder om aukande tal svakfødde kalvar. Desse kalvane hadde fellestrekk som hengande augelok (ptosis), lærevanskar, til dømes problem med å læra å drikka, mange døydde kort tid etter at dei var fødde, og dei som klarte seg vaks sakte og vart gjerne utrangert før påsett. I 2014 publiserte finske og tyske forskarar ein artikkel om desse kalvane og kalte sjukdomen PIRM-syndromet (etter symptoma «ptosis, intellectual disability, retarded growth and mortality»). Dei slo fast at PIRM er ein genetisk sjukdom med recessiv nedarving der dei sjuke kalvane har to kopiar av den sjukdomsbringande mutasjonen, mens bærarar av ein kopi er friske. Ved å samanlikna genotypane til sjuke kalvar med friske halvsøsken fann dei at det truleg er AH1 som gjev PIRM-syndromet. Etter vidare kartlegging med fullsekvensering av prøvane fann forskargruppa ein enkeltbasemutasjon i genet UBE3B. Mutasjonen fører til øydeleggande endringar i UBE3B-proteinet som spelar ein nøkkelrolle i utviklinga av organ og nervesystem under fosteruviklinga. Ein liknande mutasjon i UBE3B fins også hos menneske og gjev då opphav til Kaufman Oculocerebrofacial Syndrom med fleire symptom som liknar PIRM. Bærarfrekvensen blant finske ayrshireoksar i 2014 var 17 prosent, og med tilfeldig paring forventar ein då at 1 av 138 kalvar blir homozygot for AH1 og enten er dødfødd eller har PIRM.
AH1 i NRF – opphav og omfang
AH1-mutasjonen er ikkje blant SNPane på chipen som blir brukt til å genotypa NRF-dyr idag, så for å kartlegga omfanget av AH1 i NRF-oksane har me gjort to rundar med testar. I første runde brukte me genotypar frå ein SNP-chip med 780 000 SNPar (777K) og såg på området nær AH1-mutasjonen for å finna mulige bærarar. Så plukka me ut mistenkte historiske bærarar saman med dei siste årgangane av eliteoksar og gjorde ein spesifikk test (Agena) for AH1-mutasjonen på 364 oksar. Figur 1 oppsummerer resultatet av dei to testane og viser mulig nedarving av AH1 i NRF. I USA og Canada kan alle bærarar av AH1 sporast tilbake til oksen Selwood Betty’s Commander. Denne oksen blei importert til Finland, og AH1 kan ha komme inn i Finsk Ayrshire og SRB gjennom denne importen. NRF-oksen 3261 E. Skogmo (1982) var AH1-bærar, men den blei verken eliteokse eller oksefar så AH1 var knapt å finna i NRF fram til tidleg på 2000-talet. Men frå 2007 og utover kom AH1-mutasjonen inn i NRF gjennom import av oksane 22007 Sörby, 22008 K. Lens, 23004 Heisalan Ponistus og 23007 Asmo Tosikko Et. Desse fire importoksane har hatt stort gjennomslag som far, farfar og morfar til ein del NRF-oksar, og AH1-mutasjonen har dermed auka i frekvens i NRF dei siste 10 åra. Dei AH1-bærarane som har bidratt mest til NRF-populasjonen i dag er 11039 Skjelvan, 11335 Skjortorp og 11229 Oksjale. Andre bærarar som nyleg har vore eliteoksar er 11865 Rudland og 11876 Alm.
For å kartlegga omfanget i heile populasjonen har me utnytta genotypedata, som vanlegvis blir brukt til å rekna genomiske avlsverdiar, til å testa om genotypa dyr er sannsynlege AH1-bærar. Me finn at frekvensen av AH1 er låg, nær 0 prosent fram til 2005 og har auka til 4-5 prosent hos dyr som er fødde dei siste fem åra.
Effekt av AH1 på fruktbarheit og kalvehelse i NRF
Det finst litt over 520 000 registrerte kalvingar med både far og morfar blant dei 364 oksane med resultat i Agena-testen. Av desse kalvingane er 3,7 prosent dødfødslar. Dette talet er omtrent uendra dersom ein ser på kalvingar der enten far eller morfar er AH1-bærarar, men når både far og morfar er bærarar (totalt 1 226 kalvar) så aukar det til 5,2 prosent dødfødslar.
Ein rask analyse av kukontrolldata viser altså at dødfødslar aukar noko for kalvingar der både far og morfar er bærarar av AH1. Auken er likevel langt under 12,5 prosentpoeng som er forventinga dersom alle homozygote kalvar er dødfødde. Forklaringa kan vera at kalven har dødd så tidleg at det er registrert som kasting eller seint omløp, eller at opp mot 140 kalvar er fødd med PIRM. Å finna ut av dette vil kreva ein langt meir omfattande analyse av kukontrolldata, og ein masterstudent ved NMBU har sett på dette våren 2018.
Vegen vidare for AH1 i NRF
Kartlegging av AH1 i NRF viser at mutasjonen har komme inn i populasjonen siste 10 år og at bærarfrekvensen dei siste årgangane er 4-5 prosent. Bærarfrekvensen er så låg og bærarane er å finna i så få okselinjer at AH1 og PIRM per i dag er eit lite problem med rundt 150 dødfødde eller sjuke kalvar totalt sidan 2011. Men dersom AH1 får spre seg til fleire okselinjer og frekvensen aukar vil problemet kunna auka raskt. Me vil derfor ha som mål å fjerna AH1 frå NRF-populasjonen på sikt. Oksekatalogen vil framover få informasjon om bærarstatus, og me vil ta med AH1-status i vurderinga av eliteoksar og embryokviger. For å unngå redusert avlsframgang vil AH1-bærarar framleis kunna bli tatt ut som eliteoksar, dersom dei har svært høg avlsverdi eller ein svært god profil på indeksane. Kyr som er genotypa vil få informasjon om bærarstatus i løpet av hausten 2018 og i nye Geno avlsplan vil denne informasjonen bli presentert slik at det er enkelt å unngå bærar-bærar inseminasjonar.
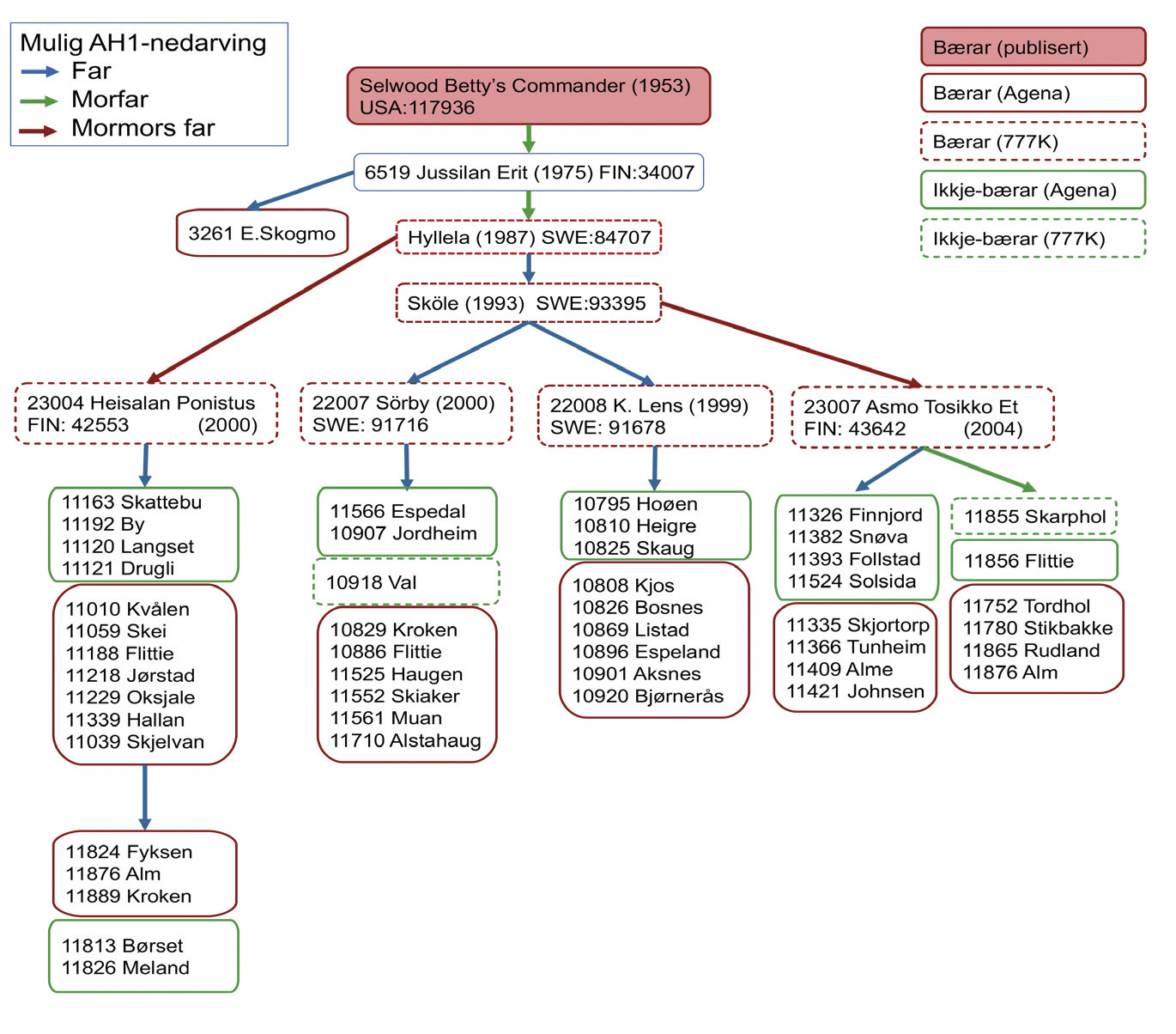
Figur 1. Oversikt over bærarstatus for NRF-oksar. Figuren gir en oversikt over bærarstatus for NRF-oksar i dei fire oksefamiliane der me finn AH1-bærarar. Figuren viser også mogeleg nedarving av AH1 via slektskap til historiske oksar frå Sverige, Finland og USA (med stamboknummer frå opphavslandet). Oksane med heiltrukken boks rundt har bærar-status bestemt med ein direkte test (Agena Biosystem MassARRAY® System) av AH1-mutasjonen, mens oksar i stipla boks har resultat frå ein noko meir usikker indirekte test (777K-genotypar i regionen rundt AH1-mutasjonen). Me manglar testresultat for den historiske oksen 6519 Jussilan Erit.
Litteraturliste kan fåast frå forfattarene.